MATURITY MOUNTAIN RAINFOREST SYNTAXA FROM SIERRA MAESTRA, EASTERN CUBA
Orlando J. Reyes1, Félix Acosta Cantillo2, Pedro Bergues Garrido3
1Ciencias Forestales
Contacto: joel@bioeco.cu
2Ing. Forestal
3Ing. Géologo
Contacto: pedro@bioeco.cu
1,2,3Eastern Center of Ecosystems and Biodiversity (BIOECO).Ministry of Science, Technology and Environment. José A. Saco Nr. 601, esq. Barnada. CP 90 100. Telf. 22626568. Santiago de Cuba, Cuba.
MODELING OF THE WASTE COLLECTION NETWORK IN THE MUNICIPALITY OF SOGAMOSO
ABSTRACT
In the Sierra Maestra the rainforests occupied mountains territories between 800 and 1 500 meters above sea level (mosl). By using the Zurich Montpellier school methodology one new order are described, Clethro cubensis- Cyrilletalia silvae with two alliances and two associations that the late and mature succesional communities are.
Keywords: Rainforests, sintaxonomy, Sierra Maestra, Eastern Cuba.
Received: May 4, 2022.Accepted: September 19, 2022
RESUMEN
En la Sierra Maestra se desarrolla la pluvisilva montana entre los 800 y los 1 500 m snm con una abundante humedad ambiental. Se estudió mediante la metodología de la Escuela Zurich Montpellier a Clethro cubensis -Cyrilletalia silvae con dos alianzas que constituyen las etapas maduras y de la sucesión avanzada.
Palabras clave: Pluvisilvas, fitocenología, Sierra Maestra, Cuba Oriental.
Recibido: 4 de mayo de 2022. Aceptado: 19 de septiembre de 2022
INTRODUCTION
Sierra Maestra has a particular paleogeographic development and its mountains are relatively old (Oro, 1989;Reyes, 2011-2012a). It also has the highest altitudes of the Cuban archipelago (up to 1 972 mosl) and has a wide variety of rainfall conditions, environmental humidity, slopes, geological conditions, soils and plant formations (Reyes, 2006; Reyes, 2011-2012b).
The vegetation of the Sierra Maestra has been relatively well studied, especially near the main roads (Reyes, 2006; Reyes, 2011-2012b). León (1946) was the first to describe the altitudinal belts of the Sierra Maestra and named them Yayales (up to 500 mosl), Manacales (500-900 mosl), Fangales (900- 1300 mosl) and Monte Fresco (1300-2000 mosl), according to the increase in altitude. Afterward Reyes (2006) makes a classification of the vegetation of the Sierra Maestra, including the montane rainforest, which is followed in this work and that was already applied in La Bayamesa National Park (Reyes &Acosta, 2005) and Pico Mogote Ecological Reserve (Reyes &Acosta, 2006) among other research. Later Reyes (2016) elaborated a forest typology based on phytocenological studies carried out with silvicultural recommendations for the restoration and / or rehabilitation of said plant formation, among others.
According to Suarez et al.(2014) the current production Sierra Maestraʹs eroded soils reaches around 50% of the potential minimum yield. The removal of biomass, where most of the nutrients are kept, as well as the elimination of litter cover and root mat (Reyes &Fornaris, 2011), minimized the possibility of nutrient recycling.
However, phytocenologic knowledge is relatively poor (Borhidi, 1996; Reyes & Acosta,2003, 2004), therefore, the objective of this work is to analyze the maturity(climax) phytocoenoses found in the montane rainforest of the Sierra Maestra, it can serve as baseline to these forest for future study about the climatic change.
METHODOLOGY
Natural conditions of the studied area
The mountain rainforest as a zonal vegetationʹs type, developed between 800 and 1 500 mosl, linking mesophyll evergreen forest beneath and cloud forest above. The soils were fundamentally allytic and ferralitic red leached (Hernandez, 2015), from deep to very deep and from acids to very acidic (ph H2O 4.0 to 5.5); the CCB (Value of S) is less than 7 cmol (+). kg-1 (Renda et al., 1981). It rains between 1 600 and 2 000 mm, with a less rainy period from November to April and with great influence of horizontal precipitation (Reyes, 2006). It were noteworthy that here between 800 and 1 200 mosl the condensation of the clouds takes place, reason why 100% of relative humidity were observed when said condensation is manifested. The relative humidity reached high values throughout the year, from just over 87% to about 92% between the different months. The average air temperature in Gran Piedra (1 100 mosl) were 18.4ºC; the minimum average was of 15.7ºC and the maximum average of 22.4ºC. As a result, evaporation were low and for eight months of the year it does not exceed 100 mm per month (Montenegro, 1990).
Sierrra Maestra divided between two parts, the Eastern Sierra Maestra (Sierra de la Gran Piedra) and the Western Sierra Maestra (Sierra del Turquino), the last were extensive and had the highest altitude.Concerning to maturity (climax) rainforests it extended sincePico Mogote (190 ,59ʹN, 75035ʹW) in the first until Pico Caracas (20001ʹN, 77002ʹW)in the last, in these terrainthe inventories were distributed.
Sampling Methodology
According to the enlarged Cuban experience, the methodology of the Zurich-Montpellier School (Braun Blanquet, 1950) were considered the most coherent with our vegetation; therefore, phytocoenological inventories (lists, stands, samples, relevés), with a minimum area of 625 m2 (Reyes, 2005) were made by this method. The abundance-dominance of each species were estimated in the usual way in this type of work: 5 when it covers 75% or more of the area of the sample; 4 covers 50 to 75%; 3 covers from 25 to 50%; 2 covers from 5 to 25%; 1 covers less than 5%; "+" with few isolated individuals and scarce coverage and "r" when they were two or three isolated individuals with limited coenological importance. In addition, observations of the oecotope were made in the place of the samples and their surroundings.
We have considered a maturity community when it herself have reached maximum development, in agreement with the local ecological conditions. At the same time, its species majority fundamentaly the main species, have a successful evolution adjusting a characteristical structure.
The ordering of vegetation inventories and the separation of phytocoenoses (syntaxa) were carried out by phytosociological methods. For the characteristic combination of the associations, the species with degrees of presence IV and V were used, and for the subassociations and variants the differential combinations (Scamoni, 1960).
In the determination of the characteristic species of the superior unit to the association, the categorization: absolutely restricted species (fidel), strongly associated and favorably associated were used (Braun Blanquet, 1950).
The Phytosociological Nomenclature International Code were followed for the categorization and nomenclature of phytocoenoses ranks (Theurillat, 2020). Completed scientific names (genus, species and author) could be observed in tables and Acevedo-Rodriguez& Strong(2012), actualized withother authors (Greuter & Rankin Rodríguez, 2016, 2017; Borhidi et al., 2017 and Sánchez, 2017,2021). Collected specimens were in Herbarium BSC.
RESULTS
According to the results obtained, the following phytosociological arrangement were made:
- Clethro cubensis-Cyrilletea silvae Reyes 2019.
(Holotypus: Magnolio cubensis-Cyrilletalia silvae Reyes2019)
This phytocoenoses occurs as a zonal formation between 800 and 2 000 m above sea level in the Sierra Maestra. It includes the mountain rainforest, the cloud forest and its advanced successional stages; it is very rich in ferns, mosses and liverworts. The soil were ferralitic red and / or yellowish leached on rocks of the El Cobre group, rainfall varied around 1 600 to 2 000 mm, and were important that it hadaround 200 days a year with fogs and low clouds.
Character species. Strongly associated: Cyrilla silvae, Magnolia cubensis subsp. cubensis, Clethra cubensis, Lyonia latifolia, Lyonia elliptica, Purdiaea stenopetala, Miconia pteroclada, Persea hypoleuca, Myrcia fenzliana, Cyathea furfuracea, Ixora ferrea, Viburnun villosum, Clusia tetrastigma, Clusia clusioides, Brunellia comocladifolia subsp. cubensis, Zeugites americanus;favorably associated: Guatteria moralesii, Palicourea alpina, Meriania albiflora, Miconia umbellata, Graffenrieda rufescens,Aiouea montana, Hedyosmum grisebachi and Arthrostylidium multispicatum.
Two orders were found:
- Magnolio cubensis-Cyrilletalia silvae Reyes2019.
- Clethro cubensis-Magnolietalia cubensis Reyes ord. nov.
In this work only the second one were studied, since the first corresponds to the cloud forest.
- Clethro cubensis-Magnolietalia cubensis Reyes ord. nov.In this contribution.
(Holotypus: Cyatheo furfuraceae-Magnolion cubensis all. nov.).
With the characteristics of the class. Although located between 800 and 1 500 mosl.Found alliances:
- Myrcio fenzlianae-Ocoteion Reyes all. nov.
- Cyatheo furfuraceae-Magnolion cubensis Reyes all. nov.
They were found in the Sierra de la Gran Piedra and the Sierra del Turquino respectively.
- Myrcio fenzlianae-Ocoteion Reyes all. nov.In this contribution.
(Holotypus: Myrcio fenzlianae-Ocoteetum cuneatae ass. nov.)
The plant formation were the mountain rainforest and its advanced successional stages; were very rich in ferns, mosses and liverworts. The soil were ferralitic red leached on rocks of the El Cobre group, the rains were around 1 600 mm and approximately 200 days with fogs and low clouds. It occurs between 800 and 1 200 mosl in the Eastern Sierra Maestra (Sierra de la Gran Piedra).
Character species.Strongly associated: Lyonia latifolia, Ocotea cuneata, Beilschmiedia pendula, Myrcia fenzliana, Clusia tetrastigma;favorably associated: Alchornea latifolia, Cyrilla silvae, Hedyosmum grisebachii, Meriania albiflora,Palicourea alpina, Brunellia comocladifolia subsp. cubensis, Psychotria grandis, Gesneria viridiflora, Cyathea aspera, Callicarpa spp., Clethra cubensis and Columnea sanguinea.
Studied association: Myrcio fenzlianae-Ocoteetum cuneatae.
- Myrcio fenzlianae-Ocoteetum cuneatae Reyes&Acosta ass. nov.In this contribution.
(Table 1, holotypus rel. 7).
This phytocoenoses with an average of 41,3 species constitutes a maturity(climax) mountain rainforest. The height of the canopy layer and its coverage were relatively uniform; in the first case it fluctuates between 12 and 15 m, while the second one between 80 and 100% of the surface. The average of species among the inventories in said layer were 15,5. The constants and most abundant were Ocotea leucoxylon(Sw.) Laness., O. cuneata(Griseb.) M. Gómez, Prestoea acuminata subsp. montana(Graham) Greuter & R. Rankin, Cupania americana L., Dendropanax arboreus(L.) Decne. & Planch., Myrcia fenzlianaO. Berg(Gomidesia lindeniana), Chionanthus domingensisLam., Matayba oppositifolia(A. Rich.) Britton, Ilex sp., Clusia tetrastigmaVesque, Syzygium jambos (L.) Alstonand Beilschmiedia pendula(Sw.) Hemsl., Cyrilla silvaeBerazaín,Aioueamontana (Sw.) R. Rohde, Alchornea latifoliaSw. and Prunus occidentalisSw. were considered frequent. Among the least frequent were Guatteria moralesii(M. Gómez) Urb.,Morella cerifera (L.) Smalland Tabebuia hypoleuca(C. Wright) Urb. Casual were Prunus myrtifolia(L.) Urb.,Cojoba arborea(L.) Britton & Rose andFicus sp. The shrub layer covers between 40 and 70%, with 10,7 species on average; the constants and sometimes abundant were Gesneria viridiflora(Decne.) Kuntze, Psychotria grandis Sw. and Cyathea parvula(Jenman) Domin. Among the frequent ones were Callicarpa oblanceolataUrb., Hedyosmum grisebachiiSolms, Meriania albifloraCarmenate & Michelang., Coccoloba diversifolia Jacq. and Cyathea aspera(L.) Sw.. The less frequent ones were represented by Casearia sylvestris subsp. myricoides(Griseb.) J. E. Gut., Lyonia latifolia(A. Rich.) Griseb.,Eugenia sp. and Miconia granulata(Urb.) Majure & Judd, while the occasional ones by Allophyllus cominia (L.) Sw. The coverage of the herbaceous layer ranges between 50 and 80%, with an average of 32,1 species. As constants were seedlings of Ocotea leucoxylon, O. cuneata, Prestoea acuminata subsp. montana, Cupania americana, Clusia tetrastigma, Chionanthus domingensisand Oplismenus hirtellus subsp. setarius (Lam.) Mez ex Ekman, Blechnum occidentaleL., Gesneria viridiflora and Psychotria grandis. Frequent species were represented by Callicarpa oblanceolata, Phaius tankervilliae(Banks) Blume, Palicourea alpina(Sw.) DC. andHedyosmum grisebachii. Less frequent were Peperomia spp., Pilea spp., Paspalum conjugatum P.J. Bergius and Homolepis glutinosa(Sw.) Zuloaga & Soderstr. The occasional ones were Prunus myrtifolia, Brunellia comocladifolia subsp. cubensisCuatrec., Cojoba arborea and Coccocypselum sp. Lianas were very poor, representing 2,4 species on average. The epiphytes, on the other hand, with 5,7 species, had in the category of constants and abundant Guzmania monostachia (L.) Rusby ex Mezand Columnea sanguinea (Pers.) Hanst.Among the less frequent were Campiloneurum phyllitidis (L.) C. Presland Bromelia sp. and as an casual Philodendron sp.
The litter were well distributed on the surface, the L layer reached between 3 and 5 cm; F and H were mixed, sometimes forming a weft that can reached up to 10 cm thick, sometimes it becomes a real root mat.
The soil were ferralitic red leached or yellowish brown, derived from rocks of the El Cobre Group; it had good internal and superficial drainage. Bioclimatically, the study area constitutes the Thermo-thermal (Humid Equatorial) type (Vilamajóet al.,1989). The Gran Piedra climatic station, adjacent to the sampled area, reported the following data (Montenegro, 1990): absolute maximum temperature 290C, absolute minimum temperature 60C, average temperature 18,40C, relative humidity 89,6%, annual average of rain 1 664 mm, average of days with rain 139, average of insolation 5 to 7 hours / day. The prevailing winds were the trade winds of the Northeast and North-northeast, which in turn were also the strongest. The average number of days with horizontal precipitations (fogs and low clouds) were 238; of them, 162 days were with dense fog.The characteristic combination can be examined in Table 1 (studied, 21-23.02.2003).
Two subassociations were observed:
- Myrcio fenzlianae-Ocoteetum cuneatae prunetosum occidentalis.
- Myrcio fenzlianae-Ocoteetum cuneatae typicum.
The differences are mainly due to the depth of the soil.
- prunetosum occidentalis Reyes subass. nov.(Table 1, typus rel. 3).
It occurs in the greatest inclinations of the association and in shallow soils. Generally, it were a greater coverage of the arboreal layer and a lower of the herbaceous one. It were a clear differential combination. There were two variants:
- Typica.
- Cojoba arborea.
The first one were characterized negatively, since several species that make up the differential combination of the second were missing. This in the variant Cojoba arborea were well defined (Table 1), missing in it Phaius tankervilliae that were part of the characteristic combination.
- typicumReyes subass. nov.(Table 1, typus rel. 7).
This subassociation occured in shallow and deep soils. It were characterized by the absence of several species that were found in the other phytocoenoses (Table 1). It had two variants:
- Typica.
- Coccoloba diversifolia.
The first were found in shallow soils and several species were missing, including Aiouea montana, which were part of the characteristic combination. The second occupied deep soils and had a clear differential combination.
Table 1. Myrcio fenzlianae-Ocoteetum cuneatae in Eastern Sierra Maestra.mpp- very shallow, Pp- shallow, Pr- deep, E3,2,1- covers (%).
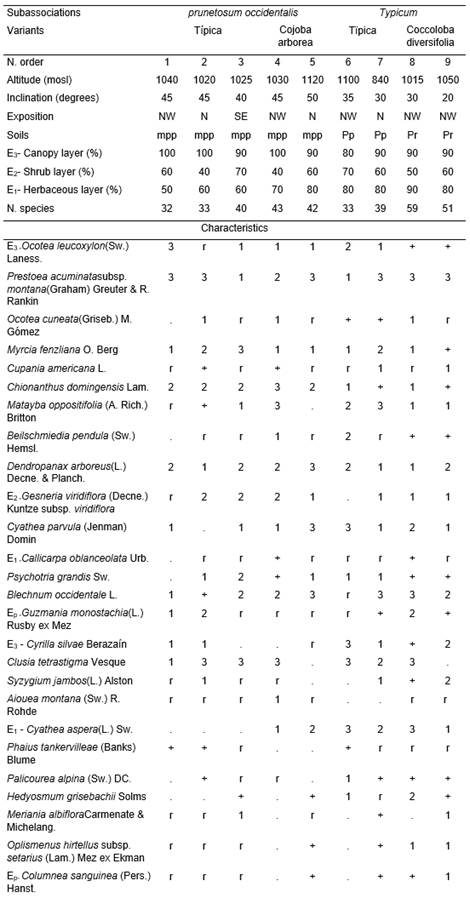
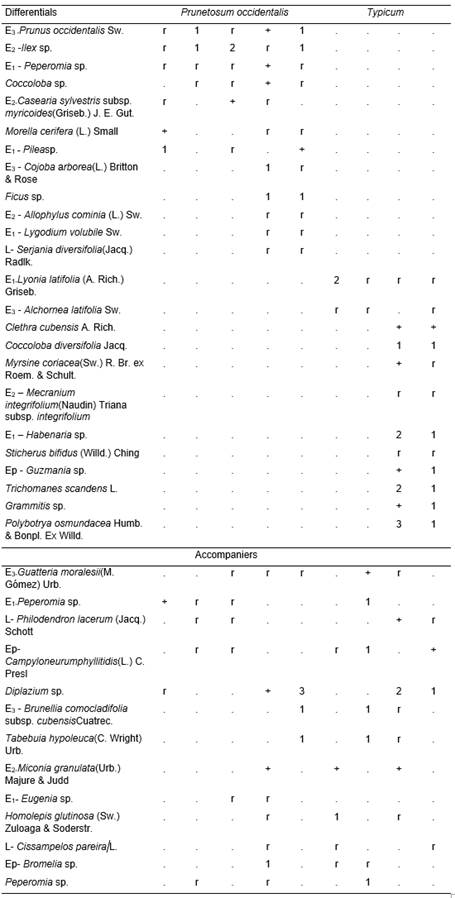
In addition.Inv. 1.Prunus myrtifolia (L.)Urb.r, Ichnanthus mayarensis (C. Wright) Hitchc. r, Paspalum conjugatum P.J. Bergius r, Marcgravia sp. r, Hohenbergia penduliflora (A. Rich.) Mez +, Phlebodium aureum (L.) J. Sm. r; Inv. 2. Tabebuiaangustata Britton r, Tillandsia flexuosa Sw. r; Inv. 3. Miconia elata (Sw.) DC. r, Pisonia aculeata L. r, Begoniasp. r, Coccocypselumsp. r, Smilaxsp. r; Inv. 4. Cyathea arborea (L.) Sm. +, Lithachne pauciflora (Sw.) P. Beauv. ex Poir. r, Canavalianitida (Cav.) Piper r, Smilax havanensis Jacq. r; Inv. 5. Cecropia peltata L. r, Myrciaria floribunda (Willd.) O. Berg r, Pavonia spinifex (L.) Cav. r, Paspalum conjugatum P.J. Bergius r, Mikania micrantha Kunth r; Inv. 6. Ilex macfadyenii (Walp.) Rehder subsp. macfadyeniir,Piper peltatum L. r, Eugenia sp. r, Scleriasp. r, Psychotria greeneanaUrb. r, Rapaneasp. r; Inv. 7. Citharexylum caudatum L. r,Scleria secans (L.) Urb. +, Mikania micrantha Kunth r, Ipomoea carolina L. r, Tillandsia fasciculata Sw. r; Inv. 8. Guettarda monocarpa Urb. +, Peperomia maculosa (L.) Hook. 1, Psychotria sp.+, Scleria lithosperma (L.) Sw. +, Coccocypselum herbaceum Aubl. r, Pseudolmedia spuria (Sw.) Griseb. r, Ipomoea carolina L. r, Isochilus linearis (Jacq.) R. Br. +, Hillia parasitica Jacq. r, Aechmea nudicaulis (L.) Griseb. r, Dichaea hystricina Rchb. f. +, Leucobrium giganteum Mull. Hal. +, Pyrrhobryum spiniformeHedw.) Mitt. +, Thuidium delicatulum(Hedw.) A.Schimp. in B.S.G.+, Blechnum fragile (Liebm.) C. V. Morton & Lellinger r, Thelypteris deltoidea(Sw.) Proctor 2; Inv. 9. Casearia sylvestris Sw. subsp.sylvestris r, Cecropia peltata L. 1, Hieronyma clusioides (Tul.) Griseb. 1, Clusia rosea Jacq. +, Ilex nitida (Vahl) Maxim. r, Miconia elata (Sw.) DC. r, Pipersp. r, Danaea nodosa (L.) Sm. +, Olyra latifolia L. 1, Oplismenus hirtellus (L.) P. Beauv. subsp. hirtellus 2, Alsophila balanocarpa(D. C. Eaton) D. S. Conant1, Sloanea curatellifolia Griseb. r, Campiloneurum sp. +.
- Cyatheo furfuraceae-Magnolion cubensis Reyes all. nov.In this contribution.
(Holotypus: Miconio pterocladae-Magnolietum cubensis ass. nov.)
This syntaxawere the maturity mountain rainforest in the Western Sierra Maestra (Sierra del Turquino) and it’s early and late successional stages, it is very rich in ferns, mosses and liverworts.It occurs between 800 and 1 500 mosl. The soil is ferralitic red leached over rocks of the El Cobre Group, the rains were around 2 000 mm and with great influence of the mists and low clouds.
Character species.Strongly associated: Magnolia cubensis subsp. cubensis, Miconia pteroclada, Cyathea furfuracea, Ixora ferrea, Guatteria moralesii, Persea hypoleuca, Tabebuia brooksiana, Zeugites americanus, Viburnum villosum, Miconia granulata, Clusia clusioides,favorably associated:Aiouea montana, Myrsine coriacea, Clethra cubensis, Miconia dodecandra, Cyrilla silvae, Prunus occidentalis, Eugenia laeteviridis, Meriania albiflora, Miconia umbellata, Palicourea alpina, Graffenrieda rufescens, Psychotria grandis, Begonia cubensis,Homolepis glutinosa and Hymenophyllum spp.
Studied association in this paper: Miconio pterocladae-Magnolietum cubensis.
- Miconio pterocladae-Magnolietum cubensis Reyes & Acosta ass. nov.In this contribution.
(Table 2, holotypus rel. 5).
This phytocoenoses with an average of 48,7 species were very complex, since it constitutes the late communities (Homeostasis II) and the maturity stage of this mountain rainforest. In its climax stage it generally had an canopy layer between 18 and 20 m, often up to 25 m or with emergent up to that height; the coverage varied between 90 and 100% and the average number of species were 14,8. The most frequent species were Magnolia cubensis Urb. subsp. cubensis, Matayba oppositifolia, Chionanthus domingensis, Prunus occidentalis, Ocotea leucoxylon, Persea hypoleuca(A. Rich.) Mez, Ixora ferrea(J.F. Gmel.) Benth., Guatteria moralesii,Aiouea montana, Miconia pterocladaUrb., Clusia clusioides(Griseb.) D’Arcy,among others. The shrub layer covered more assiduously between 40 and 60%, occasionally more or less. It had 15,6 species on average, the most abundant were Meriania albiflora, Graffenrieda rufescensBritton & P. Wilson, Palicourea alpina and Psychotria grandis; Eugenia laeteviridis Urb. and Alsophylla major Caluff & Sheltonwere also frequent. The herbaceous layer, with an average of 26,3 species, were dense, generally covered between 50 and 80%. The most abundant species vary locally and were Zeugites americanusWilld., Arthrostylidium multispicatumPilg., Peperomia maculosa(L.) Hook., Antirhea sp., Danaea ellipticaSm., Hedyosmum grisebachii and Elaphoglossum chartaceum(Baker ex Jenman) C. Chr.
This vegetation were poor in lianas, with 3,3 on average, the most found were Odontosoria aculeata (L.) J. Sm. and Arthrostylidium multispicatum. The epiphytes had an average of 7,1 species, the most frequent were Guzmania sp., Tillandsia fasciculataSw., Isochilos linearis(Jacq.) R. Br., Polybotria osmundacea Humb. & Bonpl.ex Willd. andHymenophyllum spp.
The L layer were between 1,5 and 3 cm thick, while the F generally varied between 2 and 2,5 cm; in turn, the root mat fluctuated between 6 and 13 cm, sometimes up to 20 cm in micro-depressions.
The characteristic combination had 31 species (studied 20.03.2003). There were two subassociations:
- Miconio pterocladae-Magnolietum cubensis alchorneetosum latifoliae.
- Miconio pterocladae-Magnolietum cubensis tabebuietosum booksianae.
They presented local and ecological differences.
- alchorneetosum latifoliae Reyes subass. nov.(Table 2, typus rel. 5)
It occurs at a lower altitude in the La Plata River basin and had two variants:
- Pinus maestrensis.
- Asplenium diplosceum.
The Pinus maestrensis variant occurred in the fluvial terraces of the west arm of the La Plata River and constituted a stage of Fiera II - Homeostasis II of the succession in this rainforest with the participation of Pinus maestrensisBisse. Currently said pine formed a closed canopy sublayer of around 30 to 35 m in height; in addition, there were a second sublayer between 10 and 16 m and 50 to 60% coverage, composed exclusively of broadleaf trees belonging to the rainforests. In this sublayer the most abundant species were Chionanthus domingensis (the most abundant), Miconia dodecandra(Desr.) Cogn., M. pteroclada, Myrsine coriacea, Alchornea latifolia, Aiouea montana and Ixora ferrea. The shrub layer covered between 50 and 70%, the most abundant species were Chionanthus domingensis, Viburnum villosumSw., Palicourea alpina, Ixora ferrea, Myrsine coriacea(Sw.) R. Br. ex Roem. &Schult., Meriania albiflora, Miconia dodecandra and Cyathea parvula. The herbaceous layer had between 40 and 100% coverage, the most prolific species were Palicourea alpina, Ixora ferrea, Zeugites americanus, Begonia cubensisHassk., Callicarpa ferrugineaand Meriania albiflora; there were also another large number of species. The variant Asplenium diplosceumwere the climax stage of the same rainforest.
- tabebuietosum booksianae Reyes subass nov.(Table 2, typus rel. 7)
It were located at higher altitude and on the slope of the New World stream, it had two variants:
- Danaea elliptica..
- Typica.
The first variant were the well developed rainforest, mature or climax. In turn, the Typical variant were the successional stage Homeostasis II with the participation only of broadleaf species; in it, the trees of the mountain rainforest form an canopy layer with 100% coverage and a height between 15 and 18 m with emergent relicts of around 25 m. Although young, the ecosystem equilibrium were already observed, since there were a stability of the species of this type of forest. The most abundant species were Chionanthus domingensis, Guatteria moralesii, Matayba oppositifolia, Cyrilla silvae, Clusia clusioides,Aioueamontana, Persea hypoleuca and other dispersed species. The shrub layer had 70% coverage, Cyathea furfuraceaBaker, C. parvula, Psychotria grandis and Palicourea alpina were abundant. The herbaceous layer were very diverse and covered 70%, the most abundant being Chionanthus domingensis, Guapira sp., Ocotea leucoxylon, Psychotria grandis, Palicourea alpina, Pilea sp., Homolepis glutinosa and Arthrostylidium multispicatum (Table 2).
In this stage there were a good development of the humus layers, the L layer were 3 cm, the F 1,5 cm and the H were constituted in a root mat embeded in a humus matrix of about 3 cm. The soil were leached red ferralitic, with rainfall around 2 000 mm and developed between 800 and 1 500 mosl, therefore it had fogs and low clouds most of the days of the year.
Table 2. Miconio pterocladae-Magnolietum cubensis in Western Sierra Maestra.ond – undulate.
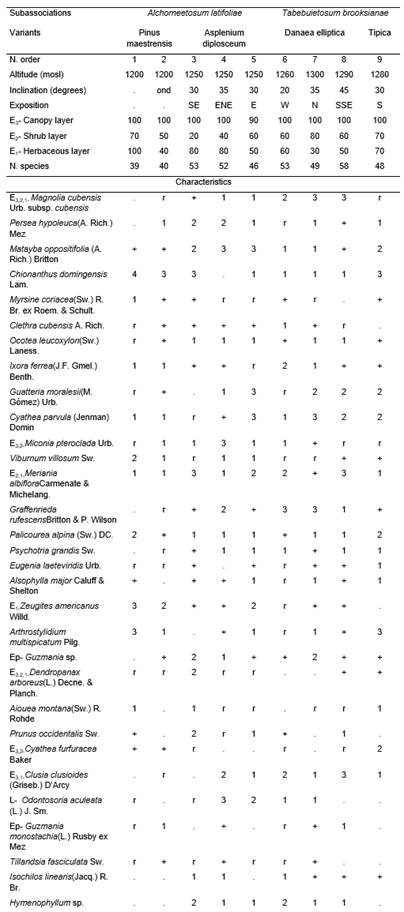

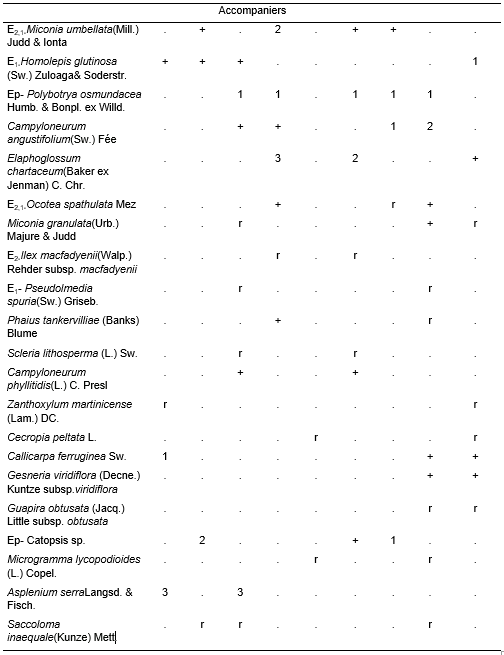
In addition.Inv. 1.Lobelia assurgens L. +, Mikania ranunculifolia A. Rich.r, Hypolepis nigrescens Hook. r; Inv. 2. Miconia norlindii (Urb.)Majure & Judd +, Antirhea sp. +, Lycopodium clavatum L. 1, Pteridium arachnoideum(Kaulf.) Maxon +, Epidendrum umbellatum Sw. +. Passiflora sexflora Juss.r; Inv. 3. Cestrum sp. r, Hirtella triandra Sw. r, Piper sp. r, Avenaria sp. r, Tillandsia sp. r, Passiflora pendulifloraBertero ex DC.r, Ipomoea tiliacea(Willd.) Choisy r, Rhipsalis baccifera (Sol. ex J. S. Muell.)Stearnsubsp. baccifera 1, Asplenium juglandifolium Lam. r; Inv. 4.Brunellia comocladifolia subsp. cubensisCuatrec. r, Phlebodium aureum (L.) J. Sm. r, Pimenta sp. r, Trophis racemosa (L.) Urb. r, Smilax domingensis Willd. r, Psychotria greeneanaUrb. r, Diplazium hastile(Christ) C. Chr.1; Inv. 5. Hieronyma nipensis Urb. r, Ichnanthussp. +, Oplismenussp. +; Inv. 6. Ilexsp. r.1, Politrichum sp. 2, Alectoria sp. 1, Mikania micrantha Kunth 1, Marcgravia sp. r; Inv. 7. Manilkara valenzuelana(A. Rich.) T.D. Penn.1, Drypetes alba Poit. +, Arachniodes denticulata(Sw.) Ching 1, Ilex sp. r; Inv. 8. Pera bumeliifolia Griseb. 1, Ilex sp. r, Ditta myricoides Griseb. r, Eupatoriumsp. r.1, Abrodictyum rigidum (Sw.) Ebihara & Dubuisson r, Dichaea glauca (Sw.) Lindl. r, Pleurothallissp. r; Inv. 9. Coccolobasp. r, Guettarda valenzuelana A. Rich. r, Pileasp. 2, Orchidaceae +, Piper peltatum L. r0, Dioscorea sp. r0, Tillandsia variabilis Schltdl.1, Diplazium unilobum(Poir.) Hieron. r, Blechnum occidentale L. r.
DISCUSSION
At these altitudes there are a conjunction of edapho-climatic conditions that determined the existence of the mountain rainforest with its structural characteristics and floristic composition that differs from the other altitudinal levels; there is also a root mat that intervenes in the feeding of the vegetation (Reyes& Fornaris, 2011) and that influences through the coverage in soil moisture (Herrera& Rodriguez,1988). The soil is leached red, oligotrophic and acidic ferralitic (Renda et al., 2012), there are strong winds and abundant rains, however temperature and relative insolation are generally low. High relative humidity and low evaporation arepresent; there is mists and clouds that influence most days of the year (Montenegro, 1990). These factors compensate and / or reinforced each other to create favorable conditions for plant development, constituting in addition one of the most important causes of high epiphytes developed(Reyes & Acosta, 2005).
It is noteworthy that the horizontal precipitations (fogs and low clouds) from 800 m above sea level occurred more frequently shortly before noon, so that in the evenings less solar radiation is usually received; this standardizes the environmental conditions during a great part of the day and minimizes at that moment the energetic differences that occur due to exposure. These horizontal precipitations can represent up to 50% of the rains(Samek &Travieso,1968). This were of great importance as suppliers of environmental humidity and limiting their loss in the ecosystem, reaching 100% saturation at the time of such low clouds, which differs from forests at higher altitudes (Reyes et al., 2012;Reyes& Acosta, 2019).
The prolific floristic composition is interesting, constituted for the most part by different species from those at lower altitudes and origin of their families (Reyes, 2006). It is also remarkable the dissimilarities between the Western Sierra Maestra and the Eastern Sierra Maestra, which are separated by elements of low altitude, the Sierra de Boniato and the depression of Santiago de Cuba, and that even consisted of different alliances.
Sintaxonomy. Borhidi (1996) exhibited the Ocoteo-Cyrilletea racemiflorae Borhidi 1996 class (24 characteristic species) and order to Ocoteo cuneatae-Magnolietalia cubensis Borhidi & Muñiz 1996. When compared the characteristic species of this class with Clethro cubensis- Magnolietea cubensis Reyes (23 species) are found to have 78.3% and 79.2% dissimilarity respectively. In turn, comparing Ocoteo cuneatae-Magnolietalia cubensis Borhidi & Muñiz 1996 (29 species) with the new order, Clethro cubensis-Magnolietalia cubensis (23 species), differ by 86.2%and 82.6% respectively. These important dissimilarities supported the conformation of these new phytocoenoses (Theurillat et al., 2020).
CONCLUSIONS
In general, in this maturity mountain rainforest had a characteristic edaphoclimatic conditions and one order, two alliances and two associations are described for the first time. This class included the advanced and mature successional stages.
Among the mountain rainforests of the eastern and western part of the Sierra Maestra, different alliances are observed.
CITED LITERATURE
[1] ACEVEDO-RODRIGUEZ, P. & M.T. STRONG. 2012. Catalogue of Seed Plants of the West Indies. Smithsonian Contributions to Botany 98. [en línea], [Consulta: 24 febrero 2020]. Disponible en: http://repository.si.edu/xmlui/handle/10088/17551.
[2] BORHIDI, A. 1996.Phytogeography and Vegetation Ecology of Cuba.2 Ed. Akadémiai Kiadó. Budapest. 926 p.
[3] BORHIDI, A., M. FERNÁNDEZ-ZEQUEIRA & R. OVIEDO PRIETO. 2017. Rubiáceas de Cuba. [en línea]. 2 Ed. S.l.: Akademiai Kiadó. Disponible en: https://go.gale.com/ps/anonymous?id=GALE%7CA581941449&sid=googleScholar&v=2.1&it=r&linkaccess=abs&issn=02366495&p=AONE&sw=w. (Consulta: Mayo 2020).
[4] BRAUN BLANQUET, J. 1950. Sociolog¡a vegetal: Estudio de las comunidades vegetales. [en línea]. Buenos Aires: Acme Agency. Disponible en: https://www.worldcat.org/title/sociologia-vegetal-estudio-de-las-comunidades-vegetales/oclc/55448784?referer=di&ht=edition. (Consulta: Junio 2019).
[5] GREUTER, W. &RANKIN RODRÍGUEZ, R. 2016. The Spermatophyta of Cuba.[en línea]. S.l.: Botanischer Garten & Botanisches Museum Berlin-Dahlem - Jardín Botánico Nacional, Universidad de La Habana. ISBN 978-3-946292-06-7. Disponible en: https://www.researchgate.net/publication/301326599. (Consulta: Junio 2017).(Consulta: Junio 2019).
[6] GREUTER, W. & R. RANKIN RODRÍGUEZ. 2017. Vascular plants of Cuba. [en línea]. Segunda edición, actualizada, de Espermatófitos de Cuba con inclusión de los Pteridófitos. S.l.: Botanischer Garten & Botanisches Museum Berlin - Jardín Botánico Nacional, Universidad de La Habana. Disponible en: https://www.bgbm.org/sites/default/files/plantas_vasculares_de_cuba_2017-12-18.pdf. (Consulta: Junio 2021).
[7] HERNÁNDEZ A., PÉREZ JIMÉNEZ J.M, BOSH D. & CASTRO SPECK N. 2015. Clasificación de los Suelos de Cuba 2015. La Habana Cuba: Instituto Nacional Ciencias Agrícolas & Instituto Suelos.[en línea]. Disponible en: http://ediciones.inca.edu.cu/files/libros/clasificacionsueloscuba_%202015.pdf.(Consulta: Junio 2018).
[8] HERRERA, R.A. & M.E. RODRÍGUEZ. 1988. Clasificación funcional de los bosques tropicales. En Herrera, R.A., L. Menéndez, M.A. Rodríguez & E.E García (Eds.). p. 574-626. Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Montevideo. ROSTLAC.
[9] LEÓN (HNO.). 1946. Flora de Cuba. Vol. 1. Contr. Ocas. Mus. Hist. Nat. del Colegio de la Salle Nr. 8, Habana. 441 p.
[10] MONTENEGRO, U. 1990. Informe con los datos climáticos de la Gran Piedra. Dpto. Meteorología de Santiago de Cuba. Academia de Ciencias de Cuba (Inédito).
[11] ORO, J.R. (1989). Evolución paleo geológica. En Nuevo Atlas Nacional de Cuba. pp. III.1.4.
[12] RENDA, A., E. CALZADILLA, J.A. BOUZA, J. ARIAS & M. VALLE. 1981. Estudio edafológico, fisiográfico y agrisilvicultural de la Sierra Maestra, Municipio Guisa. Provincia Granma. Ministerio de la Agricultura, Centro de Investigación Forestal. 90 p.
[11] RENDA, A., Y. RODRÍGUEZ, T. PLASENCIA & J. HERRERO. 2012. Manual para la Protección de los Recursos Hídricos de Cuba. Ministerio de la Agricultura. Dirección Nacional Forestal. 160 p. ISBN-978-959-7210-36-8.
[12] REYES, O.J. 2005. Estudio sinecológico de las pluvisilvas submontanas sobre rocas del complejo metamórfico. Foresta Veracruzana 7(2): 15-22.
[13] REYES, O.J. 2006. Clasificación de la vegetación de la Sierra Maestra. Editorial Academia. Biodiversidad de Cuba Oriental, Vol. VIII. p. 23–41.
[12] REYES, O.J. (2011-2012a). Zonas emergidas en Cuba Oriental, su influencia en la flora cubana. Revista Jardín Botánico Nacional 32-33: 73-78. ISSN 0253-5696.JSTOR
[13] REYES, O.J. 2011-2012b. Clasificación de la vegetación de la Región Oriental de Cuba. RevistaJardín Botánico Nacional 32-33: 59-71. ISSN 0253-5696.JSTOR
[14] REYES, O.J. 2016.Forest typology of broadleaf forests from Sierra Maestra, Eastern Cuba.Lazaroa37: 43-103. http://dx.doi.org/10.5209/LAZAROA.51054
[15] REYES, O.J. & F. ACOSTA CANTILLO. 2003. Fitocenosis presentes en las áreas costeras del Sur de la Sierra Maestra. I. Comunidades con influencia marina. Foresta Veracruzana 5(2): 1-8.
[16] REYES, O.J. & F. ACOSTA CANTILLO. 2004. Fitocenosis presentes en las áreas costeras del sur de la Sierra Maestra. II. Órdenes Varronio-Phyllostyletalia y Rhytidophyllo-Plumerietalia. Foresta Veracruzana 6(1): 37-43.
[17] REYES, O.J. & F. ACOSTA CANTILLO. 2005. Vegetation. La Bayamesa National Park, Cuba. In Rapid Biological Inventories: 13. Maceira, D., A. Fong, W.S. Alverson y T. Wachter (Eds.). p. 43–50.The Field Museum, Chicago.www.fieldmuseum.org
[18] REYES, O.J. & F. ACOSTA CANTILLO. 2006. Vegetación. Pico Mogote. In Rapid Biological Inventories.Report 09.Maceira, D., A. Fong & W.S. Alverson (Eds.). p. 40-46. The Field Museum, Chicago.www.fieldmuseum.org
[19] REYES, O.J. & F. ACOSTA CANTILLO. 2019. Bosque nublado, sus fitocenosis y vulnerabilidades ante el cambio climático. Revista Forestal Baracoa 38(1): 65-74. ISSN: 0138-6441
[20] REYES, O.J. & E. FORNARIS. 2011. Características funcionales de los bosques tropicales de Cuba Oriental. Polibotánica 32: 89-105. http://www.polibotanica.mx
[21] REYES, O.J., E. PORTUONDO, E. VADEL & E. FORNARIS. 2011. Características fisonómicas y funcionales del bosque nublado de la Sierra Maestra. Moscosoa 17: 139-149.
[22] SAMEK, V. & A. TRAVIESO. 1968. Climaregiones de Cuba. Rev. Agricultura 2: 5-23.
[23]SÁNCHEZ, C. 2017. Lista de los helechos y licófitos de Cuba. Brittonia, DOI 10.1007/s12228 017-9485-1. ISSN: 0007-196X (print) ISSN: 1938-436X (electronic, published online 23 June 2017). 24 p.
[24] SÁNCHEZ, C. 2021. Inventario de los licófitos y helechos de Cuba: sinonimia, distribución y estado de conservación. Revista del Jardín Botánico Nacional 42: 1-53. (CC BY 4.0 — https://creativecommons.org/licenses/by/4.0/)
[25] SCAMONI, A. 1960.Waldgesellschaften und Waldstandorte.Akademie-Verlag. Berlin. 326 p.
[26] SUÁREZ, C., Y. MARTÍNEZ, N. CORRÍA, L. MARTÍNEZ, A.M. RAMÓN & A. HERNÁNDEZ. 2014. Situación Ambiental de la Región Especial de Desarrollo Sostenible (REDS) Sierra Maestra (inédito). Informe Órgano de Montaña Sierra Maestra. 48 p.
[27] THEURILLAT, J.P., WILLNER, W., FERNÁNDEZ-GONZÁLEZ, F., BÜLTMANN, H., ČARNI, A., GIGANTE, D., MUCINA, L. & WEBER, H. 2020. International Code of Phytosociological Nomenclature.4th edition.Appl. Veg. Sci. doi: 10.1111/avsc.12491
[28] VILAMAJÓ, D., R.P. CAPOTE, M. FERNÁNDEZ, I. ZAMORA & B. GONZÁLEZ. 1989. Mapa bioclimático de Cuba. En Nuevo Atlas Nacional de Cuba. Inst. Geografía e ICGC. p. X. 1.2-3.
RESEÑA CURRICULAR
Principal Author: Orlando J. Reyes Domínguez. Agronomist Engineer, Dr. in Forest Science (Dr. rer. silv.), Principal Professor and Principal Investigator, Honour Member of Academy of Science. He was director of research projects and masters and doctors thesis. He was publisher and participant in nationals and internationals congress.

