OPHILITIC RAINFORESTS SYNTAXA FROM ALTIPLANICIE DE NIPE, EASTERN CUBA
1,4Orlando J. Reyes, 2,4Félix Acosta-Cantillo, 3,4Pedro Bergues-Garrido
1Dr. Ciencias Forestales joel@bioeco.cu.
2Ingeniero Forestal.
3Ingeniero Geólogo. pedro@bioeco.cu.
4Centro Oriental de Ecosistemas y Biodiversidad (BIOECO). Ministerio de Ciencia, Tecnología y Medio Ambiente (CITMA). José A. Saco Nr. 601, esq. Barnada. CP 90 100. Santiago de Cuba, Cuba
ABSTRACT
Taking in account the neccesity to know the characteristics of rainforest developed over Nipeˈs plateau, the objective was to study the phytocoenoses by using Braun Blanquet method. One alliance and two associations besides their physiognomical, ecological and phytosociological characteristics are described which is too important for their floristic relations and ecological communities analysis.
Key words: Forests, phytosociology, Sagua-Baracoa.
Recibido: 28 de enero de 2022. Aceptado: 22 de abril de 2022
Received: January 25, 2021. Accepted: April 22, 2022
FITOCENOSIS EN LA PLUVISILVA SUBMONTANA SOBRE OFIOLITAS DE LA ALTIPLANICIE DE NIPE, CUBA ORIENTAL
RESUMEN
Siendo necesario conocer las características de las pluvisilvas sobre ofiolitas en la Altiplanicie de Nipe, el objetivo del trabajo fue estudiar las fitocenosis mediante el método de Braun Blanquet. Se describen por primera vez una alianza y dos asociaciones, así como sus condiciones fisionómicas, ecológicas y fitosociológicas, lo que tiene gran impacto para analizar sus relaciones florísticas y comunitarias.
Palabras claves: Bosques, fitosociología, Sagua-Baracoa.
Como citar este artículo: Reyes, O., Acosta Cantillo, F. & Bergues-Garrido, P. (2022). Ophilitic rainforests syntaxa from altiplanicie de Nipe, Eastern Cuba, 18(35), 98-109. https://doi.org/10.33571/rpolitec.v18n35a7
1. INTRODUCTION
The rainforest are the more developed forest of the Cuban Archipelago and it is located mainly in eastern part of Cuba five kinds of rainforest were described. Reyes [1] inside which an important place is occupied by the low and submontane rainforest on ophiolites (sclerophyll rainforest) restricted in Sagua-Baracoa Subregion, also known as Nipe-Sagua-Baracoa Ridge, these are found in areas where the level of rainfalls are the highest in Cuban archipelago [2].
The geographic areas [3] where this rainforest developed is one of the more ancient in Cuba, part of which remain emerged continuously since Superior Cretaceous half [4]. Therefore are shelters of the original mountain serpentinitic flora of Cuba [4, 5].
Recently rainforest phytocoenoses were described in the eastern part of this mountains [6, 7]; and for that the objective was to study, herself from Nipe’s Mountain range.
2. MATERIALS AND METHODS
Characteristics of study area
The highest points of these territories is Loma Mensura (995 m above sea level, masl). Rocks are ophiolites, or Mesozoic ultramaphytes [8]. These rainforest are developed over dark red ferritic soils. Low and submontane rainforest on ophiolites are present since Nipe`s plateau (20033ʹNorth, 75047ʹWest – west locality) until Miel’s river basin (20013ʹNorth, 74031ʹWest – east locality), the littlest area is located at Nipe`s plateau where this study took place.
Air average temperature on highest areas is between 20 to 220C. Inside territories rain is variable; in Nipe’s Mountain range is 1 200 to 1 800 mm. In Nipe’s 1 600 to 1 800 mm are evaporated [9].
Sampling Methodology
Coordinate were took in the studied area and a 5 Km radio delimited in which territory inventories were executed with the exposed characteristic.
For studied vegetal formation denomination previously exposed criterion were followed [1, 10]. The vegetation inventories (stands, samples, relevés) were done by using the Braun Blanquet`s [11] method, following the experience of various authors [6, 7, 12].
These inventories have a minimal area of 625 m2 [13] and the abundance-dominance, sociability, vertical structure, layer and synucia description were stablished [6, 14]. In addition oecotope observations were made in stands and its surroundings; in inventories place inclination of slope, exposition, altitude (masl), general relief and nano and micro relief were measure or estimated.
The ordination of vegetation inventories and separation of syntaxa and the species presence degree (constancy) is established [15]. When inventories were five or less arabic numbers were employed and roman for six or more. For the association characteristic combinations the used species are those with degree of presence IV and V [15] and for subassociations the differential combinations. The syntaxa denomination was carrying out according to International Code of Phytosociological Nomenclature [16]. In the character species designation of the alliance recommendations [11] were used as: absolutely restricted (true, fidel), strongly associated and favorably associated, and were described the class the order and its character species too [17].
In the humus layers stratification measuring (cm), the existence of roots and rootlets were registered and roots mat characteristics. Completed scientific names (genus, species and author) could can be observed in Tables and [18] sometime amended by [19, 20, 21, 22, 23]. Collected specimens are in Herbarium BSC.
3. RESULTS
In this work one alliance and two associations were described, developing at the same time the following phytosociological arrangement:
¾ Class Tabebuio dubiae - Calophylletea utilis Reyes 2021.
Holotypus: Tabebuio dubiae - Calophylletalia utilis 2021.
¾ Order Tabebuio dubiae-Calophylletalia utilis Reyes 2021.
Holotypus: Pimento odiolentis-Calophyllion utilis Reyes 2017.
In this paper the alliance studied were: Alsophyllo minoris-Ocoteion spathulatae.
¾ Alliance Alsophyllo minoris-Ocoteion spathulatae Reyes all. nov. In this contribution.
Holotypus: Alsophyllo minoris-Ocoteetum spathulatae Reyes & Acosta ass. nov.
Composition & character species. Strongly associated: Ocotea spathulata, Tabebuia angustata, Byrsonima spicata, Coccoloba shaferi, Alchornea latifolia, Terminalia nipensis, Alsophylla minor, Piper arboreum var. stamineum, Psychotria cuspidata and Peperomia obtusifolia; favorably associated: Ocotea cuneata, Magnolia cubensis ssp. cubensis, Miconia prasina, Eugenia pinetorum and Faramea occidentalis.
The vegetal formation was submontane rainforests on ophiolites (sclerophyll rainforest). Canopy layer had about 20 m tall and 90 to 100 % cover. Rainfall was between 1 500 to 1 800 mm, with a regular distribution. In areas with soil inclination ferritic dark red from shallow to deep, with no pellets and stones is developed, locally is rocky. In humidity depressions underground water level was close to surface in part of the year, where pellets stratums were formed including a ferricreet horizon. The microrelief was sometimes irregular showing cracks. This alliance was limited to Nipe’s plateau.
Alsophyllo minoris-Ocoteion spathulatae confront with Pimento odiolentis - Calophyllion utilis Reyes 2017 had a 93 and 95 % of difference between their species. At the same time with Podocarpo ekmanii-Byrsonimion orientensis Borhidi & Muñiz 1996 had obtained 85 to 94 % of dissimilarity.
Studied associations:
¾ Alsophyllo minoris-Ocoteetum spathulatae.
¾ Tabebuio augustatae-Calophylletum utilis.
¾ Alsophyllo minoris-Ocoteetum spathulatae Reyes & Acosta ass. nov. In this contribution.
Table 1, holotypus rel. 4.
The arboreal (canopy) layer was the more rich than other one studied over ophiolites rainforest, with 34.2 species as an average between the inventories. The more plentiful and constants species were: Ocotea spathulata Mez, Hieronyma nipensis Urb., Byrsonima spicata (Cav.) DC., Chionanthus domingensis Lam., Miconia dodecandra (Desv.) Cogn., Matayba domingensis (P. DC.) Radlk., Sloanea curatellifolia Griseb., Calyptronoma occidentalis (Sw.) H.E. Moore and Cyathea parvula (Jenm.) Domin, the rest of species can be found in Table 1. Due their richness the species of shrub, herbaceous layers, lianas and epiphytes are observed in Table 1 too.
An island was conformed by this syntaxon of the submoutain rainforest on the ophiolites between the pinewood of Nipe`s high plain. It can found also in the north part of the plateau where the level of rainfalls was the biggest (about 1 800 mm) and their distribution was relatively regular.
Outside inclination fluctuates between 16 and 38 degrees and the exposition is north to northwest. Microrelief was plane cracking sometimes irregular. Soil was ferritic dark red color, between less deep to deep, with no pellets and stones, locally rocky with blocks that cover 20 to 30 % of surface; drainage was excellent as internal as external. Due vegetal cover erosion was not observed, sometimes laminar erosion was produced by swept of the superior part of fallen leaves. Generally in the first 2 cm were rich in rootlets and had an scraps structure, the quantity of rootlets progressively decreased in deep at 6 cm these rootlets disappear and only thin and coarse roots were observed. L horizon (A00) was between 1.5 and 4 cm, generally not exceed 3 cm. F (A01) was between 0.5 and 2 cm, exceptionally reach 2.5 cm; rootlets were plentiful and sometimes absorb directly for the recognized material, frequently white mycelium had observed. Bellow these root mats were present imbued in an humus matrix that vary between 5 and 12 cm, rich in rootlets and roots sometimes reaching 1 cm. In nanodepressions this root mat could reach 14 cm and conform a weft of roots and rootlets with relatively less quantity of humus in the inferior part. A fallen leaves were microlocally dregged along, and as a consequence root mat was discovered, but not forming a bare root mat.
Due the smallest area occupied by this phytocoenoses, (strongly threat) isolated from other rainforest and for that with less interchange of diasporas with other areas, homogeneity was too big following in a good way Raunkiaer`s [24] law. Characteristic combination was too rich and it`s composed by 63 species (Table 1). It was studied between 20 to 25 may 1993 (20031′North, 75042′West).
Three subassociations were found:
¾ Alsophyllo minoris-Ocoteetum spathulatae byrsonimetosum parvifoliae Reyes.
¾ Alsophyllo minoris-Ocoteetum spathulatae tabebuietosum dubiae Reyes.
¾ Alsophyllo minoris-Ocoteetum spathulatae drypetetosum glomeratae Reyes.
Mainly differences are due the soil deep, altitude and hillside position.
The subassociation byrsonimetosum parvifoliae (Table 1, typus rel. 1) present in moderately and deepest soils, located in the hillside superior parts; microrelief was flat. In the humus layers L horizon was the less developed of the association, with 1.5 cm, while the rest corresponding with the done description. The less quantity of species in the shrub (18) and herbaceous (48) layers were observed here, and so on the young arboreal species inside this layers with 13 and 18 respectively; however great quantity of shrubs in the herbaceous with fourteen were found. In the differential combination Byrsonima parvifolia Alain, Ilex macfadyeni (Walp.) Rehder ssp. macfadyenii, Calophyllum rivulare Bisse, Myrcia fenzliana O. Berg, Desmodium distortum Aubl., Oeceoclades maculata (Ldl.) Ldl., Polypodium aureum L. and Hymenophyllum polianthos (Sw.) Sw were present.
The subassociation tabebuietosum dubiae (Table 1, typus rel. 4) generally occupied the big slopes with a cracking microrelief, soil was less deep to moderately deep. Commonly was found in the superior parts of hillside. The great quantity of species and arboreal young plants were found in the herbaceous layer, with 57.2 and 24 respectively. Differential combination was small, and Tabebuia dubia (Wr. ex Sauv.) Britt. ex Seibert., Smilax domingensis Willd. were observed and in less quantity Jacaranda arborea Urb. In this syntaxon two variants were present Rhynchospora pruinosa and Cordia elliptica. The first was found in the big slopes and the rockier edafotope (20 to 30 % of rocks in surface) of the association. Differential combination were composed by Cytharexylum caudatum Willd., Rhynchospora pruinosa Griseb. and Adiantum melanoleucum Willd. Cordia elliptica variant occupied north-northeast exposition, differential combination was small, and were integred by Cordia elliptica Sw. and Palicourea barbinervis DC.
Drypetetosum glomeratae (Table 1, typus rel. 10) was located in the inferior part of slopes, generally occupied the lowest altitudes, soil was moderately deep to deep and because of its topographyc position moisture was conserved. This syntaxon was characterized of some species named in last descriptions (Table 1); differential combination was small Drypetes glomerata Griseb. and Stigmaphyllon sagreanum A. Juss. Here were found the great quantity of shrub layer species, with 28, at the same time, the bush value in the herbaceous layer was the lower, with 11.6.
Table 1. Alsophyllo minoris-Ocoteetum spathulatae in the ophiolitic rainforests of Altiplanicie de Nipe. NW – northwest, NNW – north-northwest. P - deep, Mp - moderate deep, Pp - shallow.
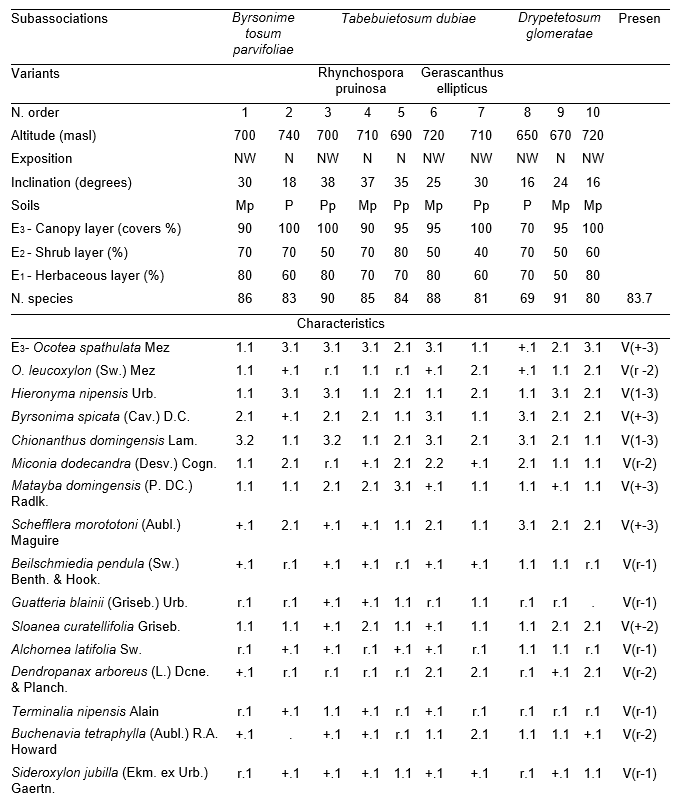
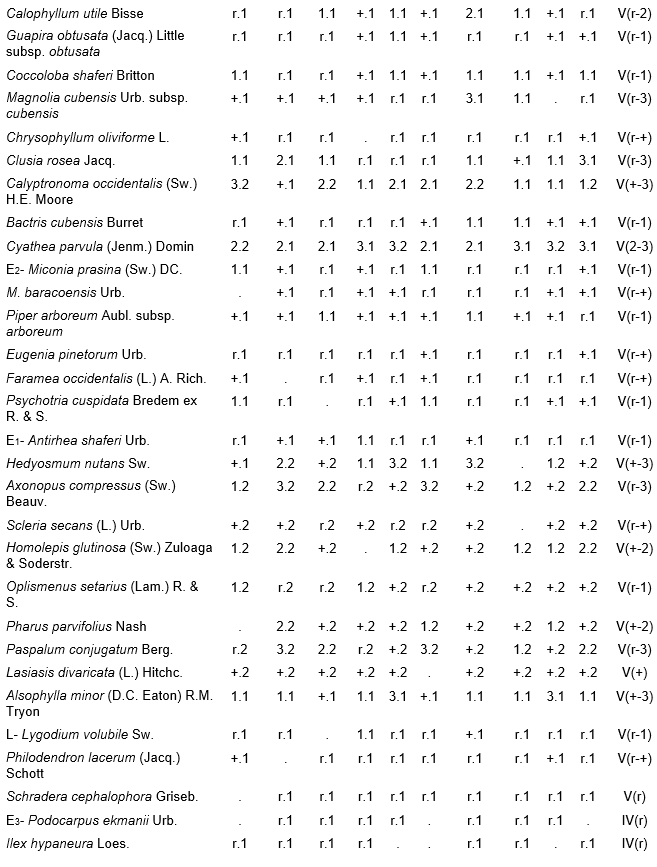
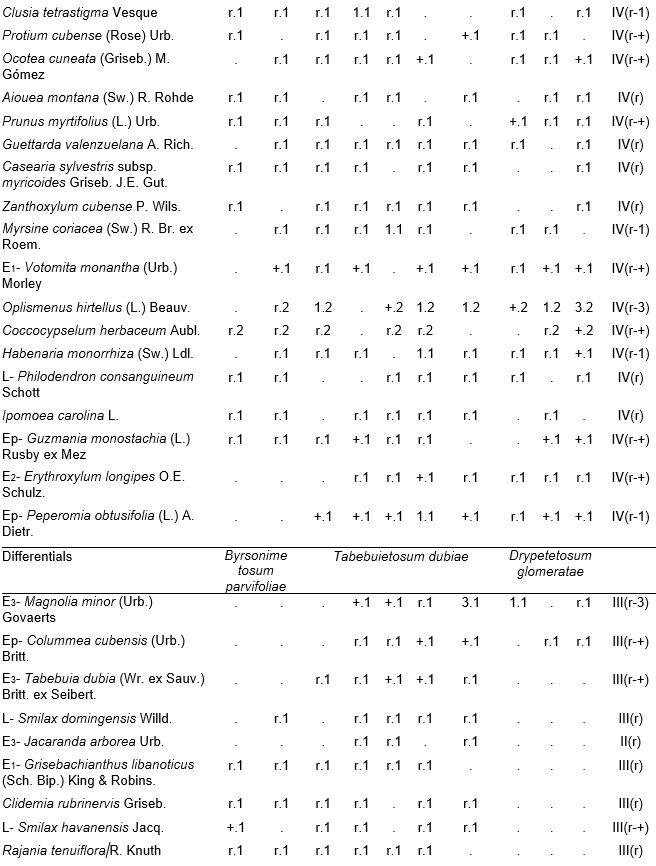
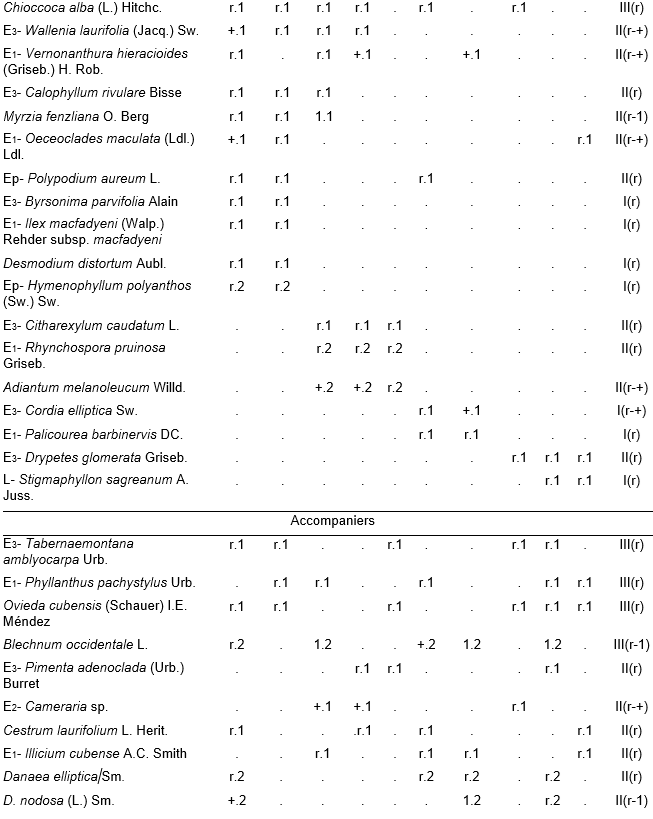
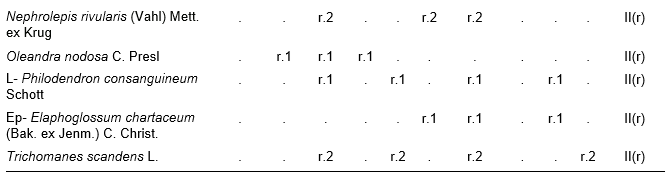
In addition. Inv. 1. Ficus velutina H. & B. ex Willd. r.1, Colubrina nipensis M.C. Johnst r.1, Nephrolepis brownii (Desv.) Hovenk. & Miyam. r.2, Securidaca elliptica Turcz r.1, Pilea microphylla Liebm. r.1, Abrus precatorius L. r.1, Ipomoea tiliacea (Willd.) Choissy r.1, Passiflora penduliflora Bert. r.1, Marcgravia evenia Krug. & Urb. r.1, Stigmaphyllon ledifolium K. Samal r.1, Clidemia sp. r.1; Inv. 2. Trema micrantha (L.) Blume r.1, Manilkara albescens (Griseb.) Cronquist r.1, Nephrolepis sp. r.2, Antirhea urbaniana C.T. White r.1, Elaphoglossum palmieri Und. & Maxon r.1, Passiflora sp. r.1, Orchidaceae r.1; Inv. 3. Tillandsia fasciculata Sw. r.1, Nephrolepis brownii (Desv.) Hovenk. & Miyam. r.2, Platygyna leonis Alain r.1, Banisteria laurifolia L. r.1, Forsteronia corymbosa (Jacq.) C.F.W. Meyer r.1, Elaphoglossum palmieri Und. & Maxon r.1, Mikania sp. r.1, Microgramma lycopodioides (L.) Copel. r.1, Hymenophyllum hirsutum (L.) Sw. r.2, Clidemia sp. r.1, Lundinia plumbea (Griseb.) B. Nord. r.1; Inv. 4. Colubrina nipensis M.C. Johnst. r.1, Pseudolmedia spuria Griseb. r.1, Elaphoglossum maxoni Underw. ex C.V. Morton r.1, Catopsis floribunda (Brongn.) L. Smith r.1, Andira jamaicensis (Wr.) Urb. r.1, Guapira sp. r.1, Eugenia sp. r.1, Hymenophyllaceae r.2; Inv. 5. Passiflora sexflora A. Juss. r.1, Vanilla palmarum Salzman ex Lindl. r.1, Platygyna leonis Alain r.1, Karwinskia bicolor (Britt. & Wils.) Urb. r.1, Polygala oblongata (Britt.) Blake r.1, Lunania dodecandra Wr. r.1, Catesbaea spinosa L. r.1, Pseudolmedia spuria Griseb. r.1, Commelina sp. r.1, Orchidaceae r.2; Inv. 6. Tillandsia fasciculata Sw. r.1, Tournefortia hirsutissima L. r.1, Asketanthera calycosa (A. Rich.) Woods r.1, Solanum torvum Sw. r.1, Passiflora sexflora A. Juss. r.1, Calyptranthes subcapitata Urb. r.1, Olfersia alata C. Sanchez & Caluff r.2, Psychotria grandis Sw. r.1, Vanilla palmarum Salzman ex Lindl. r.1, Catopsis floribunda (Brongn.) L. Smith r.1, Forsteronia corymbosa (Jacq.) C.F.W. Meyer r.1, Solanum pachyneurum O.E. Schulz r.1; Inv. 7. Ficus velutina H. & B. ex Willd. r.1, Triopteris rigida Sw. r.1, Rhamnidium ellipticum Britt. & Wils. r.1, Prunus occidentalis Sw. r.1, Sapium adenodon Griseb. r.1, Hyperbaena domingensis (DC.) Benth. r.1, Antirhea urbaniana C.T. White r.1; Inv. 8. Hyperbaena domingensis (DC.) Benth. r.1, Brunfelsia cestroides A. Rich. r.1, Pecluma pectinata (L.) M.G. Price r.1, Euphorbiaceae r.1, Ouratea striata (V. Tiegh) Urb. r.1, Mouriri purpurascens Urb. r.1, Psychotria grandis Sw. r.1, Simaruba laevis Griseb. r.1, Prunus occidentalis Sw. r.1, Tillandsia bulbosa Hook. r.1, Orchidaceae r.1; Inv. 9. Brunfelsia cestroides A. Rich. r.1, Turpinia paniculata Vent. r.1, Serpocaulon loriceum (L.) A.R. Sm. r.1, Catopsis berteroniana (Schult.) Mez r.1, Pera bumeliifolia Griseb. r.1, Ouratea striata (V. Tiegh) Urb. r.1, Polygala oblongata (Britt.) Blake r.1, Microgramma lycopodioides (L.) Copel. r.1, Erythroxylum sp. r.1 Orchidaceae r.2; Inv. 10. Adiantum pyramidale (L.) Willd. r.1, Catopsis berteroniana (Schult.) Mez r.1, Pimenta racemosa (Mill.) J.W. Moore r.1, Spathelia pinetorum M. Vict. r.1, Pera bumeliifolia Griseb. r.1, Erythroxylum sp. r.1, Clidemia sp. r.1.
¾ Tabebuio angustatae - Calophylletum utilis Reyes & Acosta ass. nov. In this contribution.
Holotypus: this relevé.
This isolated syntaxa was the only and little one relict of the submountain rainforest on ophiolites that was present in very wet, even marshy soils of Nipe’s high plain. That`s for it was seriously threatened. The mainly fallen leaves contributors (more than 75 %) were Calophyllum utile Bisse, Matayba domingensis and Sloanea curatellifolia. There still remain a little relict in good structural and floristic conditions, the association was characterized for a phytocoenologic relevé that represent it`s holotype.
E3 – Canopy layer, 20 m high and 90 % cover.
Calophyllum utile 4.1, Matayba domingensis 2.1, Ocotea spathulata 1.1, Sideroxylon jubilla (Ekm. ex Urb.) Gaertn. +.1, Ocotea leucoxylon (Sw.) Mez +.1, Sloanea curatellifolia +.1, Guatteria blainii (Griseb.) Urb. +.1, Schefflera morototoni (Aubl.) Maguire +.1, Dendropanax arboreus (L.) Decne. & Planch. +.1, Clusia rosea Jacq. 1.1, Buchenavia tetraphylla (Aubl.) R.A. Howard +.1, Tabebuia angustata Britton +.1, Cyrilla coriacea Berazaín 1.1.
E2 – Shrub layer, 50 % of cover (last mentioned were excluded)
Ilex macfadyenii ssp. macfadyenii +.1, Chionanthus domingensis +.1, Cyathea parvula 3.2, Tabernaemontana amblyocarpa Urb. 1.1, Bactris cubensis Burret +.1, Ocotea cuneata (Griseb.) M. Gómez r.1, Coccoloba shaferi Britton +.1, Byrsonima spicata r.1, Faramea occidentalis (L.) A. Rich. +.1, Piper arboreum Aubl. subsp. arboreum +.2, Clusia tetrastigma Vesque r.1.
E1 – Herbaceous layer, 90 % cover (last mentioned were excluded)
Arthrostylidium fimbriatum Griseb. 4.3, Coccocypselum herbaceum Aubl. 1.2, Homolepis glutinosa (Sw.) Zuloaga & Soderstr. 1.2, Myrsine coriacea (Sw.) R. Br. ex Roem. r.1, Miconia prasina (Sw.) DC. r.1, Abarema glaucum (Urb.) Barneby & J.W. Grimes r.1, Vernonanthura hieracioides (Griseb.) H. Rob. r.1, Peperomia maculosa (L.) Hook. r.1, P. obtusifolia (L.) A. Dietr., Oplismenus hirtellus (L.) Beauv. +.2, Axonopus compressus (Sw.) Beauv. 1.2, Scleria secans (L.) Urb. 1.2, Beilschmiedia pendula (Sw.) Hemsl. r.1, Clidemia cubensis Cogn. r.1, Piper sp. +.1, Nephrolepis rivularis (Vahl) Mett. ex Krug. +.2.
Lianas and climbing: Smilax domingensis +.1, Philodendron lacerum (Jacq.) Schott +.1, Chiococca alba (L.) Hitchc. r.1, Marcgravia evenia Krug. & Urb. subsp. evenia r.1, Abrus precatorius L. r.1, Schradera cephalophora Griseb. r.1, Platygyna hexandra (Jacq.) Müll. Arg. r.1, Polybotrya osmundacea Humb. & Bonpl. ex Willd. r.1.
Epiphytes: Tillandsia fasciculata Sw. +.1, T. usneoides (L.) L. +.2, Elaphoglossum chartaceum (Baker ex Jenman) C. Chr. +.2.
It′s founded in an open depression, slightly concave where the surroundings drain water is concentred. For that reason the presence of water table was close to soil surface in part of the year and directly on surface during the wet period it means that periodically was flooded. In neighboring areas in central part of the depression the substratum were conformed by pellets and ferricreet as a product of pellets consolidation; this process was produced under reduction conditions because of poor drainage. Humus layers were good developed and defined. L had a thickness between 2 and 3 cm and F for about 1 cm. H layer conformed a root mat of 10.5 cm imbued in a black humus matrix, very wet and full of roots and rootlets. Under Calophyllum utile trees F layer reach 2.5 cm and root mat 16.5 cm. Rainfall level gets about 1 500 mm, and is complemented by the edaphic wet. Coexistence of both factors supports this rainforest. It was studied on 27 may 1993 (20031′North, 75046′West).
4. DISCUSSION
Nipe’s plateau is the west limit of Sagua-Baracoa massif and them the more open geographic area to floristic interchange with other areas. Even like submountain rainforest on ophiolites of the area that make up small and isolated segments, conditioned for north exposition in a great rainfall zone of the system (Alsophyllo minoris-Ocoteetum spathulatae) and for occupy very wet depressions (Tabebuio angustatae-Calophylletum utilis), are more exposed to the introduction of elements from surroundings ecosystems. For that reasons Alsophyllo minoris-Ocoteetum spathulatae is the most prolific of all the associations (83.7 species average). At the same time, the limited area occupied for this phytocoenoses enclosed by pinewood, makes this very homogeneous and for that has the more rich characteristic combination (64 species).
When the canopy layer is analyzed, once more is observed that Alsophyllo minori-Ocoteetum spathulatae is the more prolific syntaxon with 32.4 species average.
It`s well know that in spite of these ferritic soils are acidic and poor [25] present in high slopes with no pellets have a nutrient provision and a percentage of pores and profitable water big than the lend undulating zones (occupied by pinewood) of the surrounded [26], which help the broadleaf mainly the trees. Due its strong inclination with north-northwest exposition, the occupied area by Alsophyllo minori-Ocoteetum spathulatae received less calorific energy than the adjacent areas, this conditions permit that the soil always maintain fresh with enough moisture to support exuberant vegetation. These factors locally conditioned the presence of this rainforest in a land domined by pinewood.
As exposed previously [6, 27, 28, 29], the forests that present this kind of root mat have an storage system and recycled nutrients with a great significance because of almost the totally of this recycled nutrients are produced with its own participation so in this ecosystemic function soil participation is minimized.
5. CONCLUSIONS
Alsophyllo minoris-Ocoteetum spathulatae is the west and richest phytocoenose from submontain ophilitic rainforest from Sagua Baracoa Mountains that make up small and isolated segments, conditioned for north exposition, more provision and a percentage of pores and profitable water and less calorific energy than the adjacent areas. It`s the more prolific syntaxon in the canopy layer too with 32.4 species average.
6. CITED LITERATURE
[1] Reyes O.J. Clasificación de la vegetación de la Región Oriental de Cuba. Revista Jardín Botánico Nacional 32-33: 59-71. 2011-2012a. http://www.rjbn.uh.cu/
[2] Guevara, V., Rodríguez Y. & Roque A. (Redactores Temáticos). Precipitación media hiperanual 1961-2000. En: Atlas Nacional de Cuba LX Aniversario. 2019.
[3] Núñez Jiménez A. & Viña Bayés N. Regiones Naturales Antrópicas. En: Inst. Geografía e ICGC (eds.). Nuevo Atlas Nacional de Cuba. p. XII.2.1. La Habana. 1989.
[4] Reyes O.J. Zonas emergidas en Cuba Oriental, su influencia en la flora cubana. Revista Jardín Botánico Nacional 32-33: 73-78. 2011-2012b. http://www.rjbn.uh.cu/
[5] Borhidi A. Phytogeography and Vegetation Ecology of Cuba. 2 Ed. Akadémiai Kiadó. Budapest. 1996.
[6] Reyes O.J. & Acosta Cantillo F. Fitocenosis en las pluvisilvas sobre ofiolitas del Parque Nacional Alejandro de Humboldt, Cuba Oriental. Caldasia 3(1): 91-123. 2017.
[7] Reyes O.J. & Acosta Cantillo F. Fitocenosis en la pluvisilva de baja altitud sobre rocas metamórficas, Cuba Oriental. Revista del Jardín Botánico Nacional 39: 49-57. 2018. http://www.rjbn.uh.cu/
[8] Pérez, C.M., Chuy T. & Geler T. (Redactores Temáticos). Geología. En: Atlas Nacional de Cuba LX Aniversario. 2019.
[9] Crespo S.E. Evaporación media anual. In: Inst. Geografía e ICGC (eds.). Nuevo Atlas Nacional de Cuba. Mapa 42. p. VI.4.1. La Habana. 1989.
[10] Reyes O.J. & Acosta Cantillo F. Vegetación. Cuba: Parque Nacional Alejandro de Humboldt. In: Rapid Biological Inventories 14. Maceira, D., A. Fong, W.S. Alverson & T. Wachter (Eds.). p. 54–69. 2005. www.fieldmuseum.org
[11] Braun Blanquet J. Sociología vegetal: Estudio de las comunidades vegetales. 1950. https://www.worldcat.org/title/sociologia-vegetal-estudio-de-las-comunidades-vegetales/oclc/55448784?referer=di&ht=edition.
[12] Galán de Mera A. & Vicente Orellana J.A. Aproximación al esquema sintaxonómico de la vegetación de la región del Caribe y América del Sur. Anales de Biología 28: 3-27. 2006.
[13] Reyes O.J. Estudio sinecológico de las pluvisilvas submontanas sobre rocas del complejo metamórfico. Foresta Veracruzana 7(2): 15-22. 2005.
[14] Reyes O.J. & Acosta Cantillo F. Fitocenosis en los bosques siempreverdes de Cuba Oriental. III. Pruno-Guareetum guidoniae en la Sierra Maestra. Foresta Veracruzana 13(1): 1-6. 2011.
[15] Scamoni A. & Passarge H. Einführung in die praktische Vegetationskunde. 2 Aufl. Jena. 1963. https://books.google.com.cu/books/about/Einf%C3%BChrung_in_die_praktische_Vegetation.html?id=eA51mQEACAAJ&redir_esc=y.
[16] Weber H.E., Moravec J. & Theurillat J.P. International Code of Phytosociological Nomenclature. 3rd Edition. Journal of Vegetation Science 11: 739-768. 2000. DOI 10.2307/3236580.
[17] Reyes, O.J., Acosta, F. & Bergues, P. Sierra del Cristal and Alturas de Moa its ophiolitic rainforests syntaxa. Revista Politécnica (17)34: 181-195. 2021. DOI: https://doi.org/10.33571/rpolitec.v17n34a12.
[18] Acevedo-Rodriguez P. & Strong M.T. Catalogue of Seed Plants of the West Indies. Smithsonian Contributions to Botany 98. Scholarly Press. Washington D.C. 1192 p. 2012. http://repository.si.edu/xmlui/handle/10088/17551.
[19] Greuter W. & Rankin Rodríguez R. The Spermatophyta of Cuba. A Preliminary Checklist. Part II: Checklist. Botanischer Garten - Botanisches Museum Berlin-Dahlem & Jardín Botánico Nacional, Universidad de La Habana. 398 p. 2016. DOI: http://dx.doi.org/10.3372/cubalist.2016.1
[20] Greuter W. & Rankin Rodríguez R. Vascular plants of Cuba. A preliminary checklist. Second, updated Edition of The Spermatophyte of Cuba, with Pteridophyte added. Botanischer Garten - Botanisches Museum Berlin-Dahlem & Jardín Botánico Nacional, Universidad de la Habana. 444 p. 2017. DOI: 10.3372/cubalist.2017.1
[21] Borhidi A., Fernández-Zequeira M. & Oviedo Prieto R. Rubiáceas de Cuba. Akademiai Kiadó. Budapest.2017. https://go.gale.com/ps/anonymous?id=GALE%7CA581941449&sid=googleScholar&v=2.1&it=r&linkaccess=abs&issn=02366495&p=AONE&sw=w.
[22] Sánchez C. Lista de los helechos y licófitos de Cuba. Brittonia, 2017. DOI 10.1007/s12228-017-9485-1. ISSN: 0007-196X (print) ISSN: 1938-436X. 23 June 2017.
[23] Sánchez, C. Inventario de los licófitos y helechos de Cuba: sinonimia, distribución y estado de conservación. Revista del Jardín Botánico Nacional 42: 1-53. 2021. (CC BY 4.0 — https://creativecommons.org/licenses/by/4.0/)
]24]-Raunkiaer C. The life forms of plants an statistical plant geography. Oxford: Oxford University Press. 1934.
[25] Hernández A., Pérez Jiménez J.M, Bosh D. & Castro Speck N. Clasificación de los Suelos de Cuba 2015. Instituto Suelos e Instituto Nacional de Ciencias Agrícolas. La Habana. 2015. http://ediciones.inca.edu.cu/files/libros/clasificacionsueloscuba_%202015.pdf.
[26] Reyes O.J. Untersuchungen über die Formation der Kiefernwälder in Pinares de Mayarí (Kuba) und Vorschläge zur Erhöhung ihrer Leistungsfähigkeit. [Dr. Sc. Diss.] [Dresden TU] Deutschland. 1977.
[27] Jordan, C.F. Ed. An Amazonian Rain Forest. The Structure and Function of a Nutrient Stressed Ecosystem and the Impact of Slash and Burn Agriculture. Man and Biosphere Series. Vol. 2. 176 p. 1989.
[28] Rangel-Ch, J.O. La Vegetación de la Región Amazónica de Colombia. Aproximación inicial. 1-53. En: Colombia Diversidad Biótica VII. Vegetación, Palinología y Paleoecología (Rangel-Ch, J.O, Ed.).Universidad Nacional de Colombia. Bogotá. 2008.
[29] Reyes, O.J. & Fornaris E. Características funcionales de los bosques tropicales de Cuba Oriental. Polibotánica 32: 89-105. 2011. http://dx.doi.org/10.15446/caldasia.v39n1.64581.